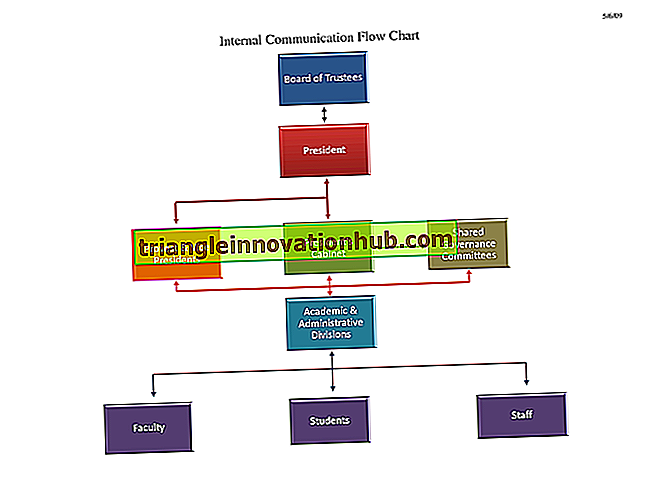
Tế bào vi khuẩn: Các bộ phận khác nhau của tế bào vi khuẩn tổng quát (được giải thích bằng sơ đồ) | Vi trùng học
Các phần khác nhau của tế bào vi khuẩn tổng quát đã được thể hiện trong Hình 2.3 và được mô tả như sau:

1. Flagella:
Flagella vi khuẩn là phần phụ xoắn ốc giống như sợi tóc mỏng nhô ra qua thành tế bào và chịu trách nhiệm cho sự di chuyển của vi khuẩn. Hầu hết các vi khuẩn vận động sở hữu Flagella.
Chiều dài của nó là khoảng 10-15. Flagella vi khuẩn hoàn toàn khác với Flagella eukaryote về cấu trúc và cơ chế hoạt động. Chúng mỏng hơn nhiều so với Flagella hoặc lông mao của các tế bào nhân chuẩn. Không có màng tế bào hiện diện trên vi khuẩn Flagella không giống như ở vi khuẩn nhân chuẩn.
Hơn nữa, vi khuẩn Flagella, không giống như vi khuẩn eukaryote, hơi cứng nhắc và không quất qua lại, thay vào đó chúng quay giống như chân vịt của một chiếc thuyền cho sự di chuyển của tế bào. Flagella vi khuẩn có ba phần là; (a) Cơ thể cơ bản, (b) Móc và (c) Dây tóc (Hình 2.4).

(a) Cơ thể cơ bản:
Nó tạo thành phần cơ sở của lá cờ và được neo trong màng tế bào chất và thành tế bào. Nó cung cấp neo cho Flagella cũng như các chức năng như một động cơ, do đó truyền đạt chuyển động quay của cột cờ.
Nó bao gồm một thanh trung tâm nhỏ đi qua một hệ thống các vòng. Trong trường hợp vi khuẩn gram dương, trong đó thành tế bào chỉ bao gồm một lớp peptidoglycan, cơ thể cơ bản chỉ có một cặp vòng; vòng M và vòng S, cả hai được nhúng trong màng tế bào.
Bao quanh các vòng này là một nhóm protein gọi là protein mot. Sự di chuyển của proton qua màng tế bào thông qua các protein mot này tạo ra lực động lực proton (năng lượng) cần thiết cho sự quay của hai vòng này, cuối cùng đóng vai trò là động cơ cho vòng quay giống như chân vịt của cột cờ.
Một nhóm protein khác, được gọi là protein fli, có mặt kẹp giữa hai vòng này. Chúng hoạt động như một công tắc động cơ, đảo ngược sự quay của lá cờ để đáp ứng với các tín hiệu nội bào.
Trong trường hợp vi khuẩn gram âm, trong đó thành tế bào bao gồm một lớp lipopolysacarit bên ngoài (LPS) bên cạnh lớp peptidoglycan bên trong mỏng hơn, cơ thể cơ sở có thêm một cặp vòng; vòng L được nhúng trong lớp LPS và vòng P trong lớp peptidoglycan mỏng.
(b) Móc:
Một cái móc ngắn nối cơ thể cơ bản với dây tóc của lá cờ. Nó hơi dày hơn dây tóc. Hành động vận động của cơ thể cơ sở được truyền đến dây tóc thông qua móc.
(c) Dây tóc:
Đó là phần sợi tóc giống như lông dài của lá cờ, nổi lên từ lưỡi câu. Nó dài hơn nhiều lần so với tế bào vi khuẩn. Nó được làm từ một loại protein duy nhất, 'flagellin' có trọng lượng phân tử khoảng 40.000. Phân tử Flagellin tạo thành chuỗi; ba chuỗi như vậy đan xen nhau để tạo thành cột cờ. Không giống như một sợi tóc mọc ở gốc của nó, dây tóc của một lá cờ mọc ở đầu của nó.
Các axit amin đi dọc theo trung tâm rỗng của dây tóc và thêm vào đầu xa của nó. Quá trình này được gọi là tự lắp ráp, vì tất cả các thông tin cho cấu trúc cuối cùng của dây tóc nằm trong chính các tiểu đơn vị protein. Dây tóc có phần cứng nhắc và không quất qua lại, thay vào đó, nó quay tròn giống như chân vịt của một chiếc thuyền cho sự di chuyển của tế bào.
Các xoắn khuẩn thiếu Flagella bên ngoài. Tuy nhiên, chúng di chuyển với sự trợ giúp của cấu trúc giống như Flagella nằm trong các tế bào của chúng ngay bên dưới lớp vỏ bên ngoài. Chúng được gọi là flagella periplasmic, flagella axial, fibrils trục hoặc endoflagella.
Flagella được sắp xếp theo những cách khác nhau trong các vi khuẩn khác nhau. Sự sắp xếp của Flagella trên các tế bào vi khuẩn được gọi là flagname. Flagname có bốn loại sau (Hình 2.5).

(a) Cờ hiệu đơn sắc (Mono: đơn; trichous: tóc): Một lá cờ đơn có mặt ở một đầu của tế bào.
(b) Lophotrichous flagname (Lopho: tuft; trichous: tóc): Một búi hoặc cụm Flagella có mặt ở một đầu của tế bào.
(c) Cờ hiệu lưỡng tính (Amphi: cả hai; trichous: tóc): Flagella có mặt ở cả hai đầu của tế bào hoặc đơn lẻ hoặc ở dạng búi.
(d) Flagit Peritrichous (Peri: xung quanh; trichous: tóc): Flagella có mặt trên khắp bề mặt của tế bào.
2. Fimbriae:
Fimbriae là những phần phụ giống như sợi tóc mỏng, có cấu trúc tương tự như Flagella, nhưng không liên quan đến sự vận động của vi khuẩn (Hình 2.3). Không giống như Flagella, chúng ngắn hơn đáng kể và nhiều hơn, nhưng giống như Flagella, chúng được làm từ các phân tử protein.
Fimbriae có mặt trong một số loài vi khuẩn, giúp chúng bám vào các tế bào của các sinh vật khác và ngăn chúng bị cuốn trôi bởi dòng chảy của chất nhầy hoặc chất lỏng cơ thể. Do đó, nó đặc biệt giúp vi khuẩn gây bệnh thiết lập sự lây nhiễm trong cơ thể vật chủ. Chúng cũng giúp hình thành các hạt hoặc màng sinh học trên bề mặt.
3. Pili:
Pili là phần phụ giống như sợi tóc mỏng, có cấu trúc tương tự như Flagella, nhưng không liên quan đến sự vận động của vi khuẩn (Hình 2.6). Chúng cũng có cấu trúc tương tự như fimbriae, nhưng thường dài hơn và chỉ có một hoặc một vài pili hiện diện trên bề mặt tế bào vi khuẩn.

Nó được làm từ một loại protein duy nhất, 'pilin', có trọng lượng phân tử khoảng 17.000. Các phân tử Pilin được sắp xếp một cách xoắn ốc để tạo thành một sợi cứng duy nhất với lõi rỗng trung tâm. Ở một số vi khuẩn, vật liệu di truyền được truyền qua lõi rỗng của pili trong quá trình liên hợp (giao phối với vi khuẩn khác).
Pili như vậy được gọi là pili sinh sản (F pili) hoặc pili sex. Ở một số vi khuẩn gây bệnh, pili hoạt động tương tự như fimbriae trong việc gắn vào các tế bào chủ và thiết lập sự lây nhiễm. Hoạt động ngưng kết của pili dẫn đến sự hình thành các màng vi khuẩn có thể nhìn thấy trên bề mặt nuôi cấy nước dùng.
4. Lớp S:
Lớp S (lớp bề mặt) là lớp được tìm thấy trên bề mặt của hầu hết các vi khuẩn gây bệnh, bao gồm một chuỗi các protein hai chiều có hình dạng tinh thể trong các đối xứng khác nhau, như hình lục giác, hình tứ giác hoặc hình tam giác (Hình 2.7) .

Sự đối xứng tinh thể phụ thuộc vào thành phần protein của lớp S. Nó được liên kết với cấu trúc thành tế bào (với lớp peptidoglycan ở vi khuẩn gram dương và với lớp LPS ở vi khuẩn gram âm).
Nó hoạt động như một hàng rào thấm bên ngoài, cho phép đi qua các chất có trọng lượng phân tử thấp và loại trừ các chất có trọng lượng phân tử cao. Nó cũng cung cấp sự bảo vệ cho các vi khuẩn gây bệnh, chống lại cơ chế bảo vệ vật chủ.
5. Viên nang (lớp chất nhờn hoặc glycocalyx):
Nhiều vi khuẩn có chất nhớt nhớt hoặc chất keo bên ngoài tế bào, bao quanh thành tế bào (Hình 2.8). Nó được làm từ các đại phân tử, không liên kết cộng hóa trị với vi khuẩn.

Các đại phân tử bề mặt này chủ yếu là các polysacarit tích điện (glycoprotein hoặc glycolipids) và do đó được gọi là 'glycocalyx' (glyco: carbohydrate; calyx: một phần của hoa bên dưới cánh hoa kéo dài ra khỏi cuống). Glycocalyx được định nghĩa là vật liệu chứa polysacarit nằm bên ngoài tế bào như một mạng lưới lỏng lẻo của các sợi polymer kéo dài ra khỏi tế bào.
Chúng được tổng hợp và tiết ra bởi các vi khuẩn, liên kết với các thụ thể trên bề mặt của các mô chủ. Glycocalyx không chỉ được tìm thấy xung quanh một số tế bào vi khuẩn; nó cũng được tìm thấy xung quanh một số tế bào thực vật và động vật.
Glycocalyx có thể có ba loại như sau:
(a) Viên nang:
Nó là một lớp phủ polymer bao gồm một lớp nhớt dày đặc, được xác định rõ ràng bao quanh các tế bào vi khuẩn ở nhiều loài. Lớp nhớt có thể được hiển thị bằng kính hiển vi ánh sáng bằng kỹ thuật nhuộm màu đặc biệt.
Các sợi polymer glycocalyx được tổ chức trong một ma trận chặt chẽ loại trừ các hạt như mực Ấn Độ. Vi khuẩn, có nang, được gọi là vi khuẩn bọc, trong khi những vi khuẩn không có nang, được gọi là vi khuẩn không bọc.
(b) Vi nang:
Đó là một lớp nhớt bao quanh các tế bào vi khuẩn, mỏng đến mức không thể nhìn thấy bằng kính hiển vi ánh sáng.
(c) Chất nhờn:
Nó là một khối khuếch tán của các sợi polymer dường như không liên kết với bất kỳ tế bào đơn lẻ nào. Lớp nhớt dày đến mức nhiều tế bào được nhúng trong ma trận chất nhờn. Lớp dễ bị biến dạng và không loại trừ các hạt. Rất khó để được hình dung bằng kính hiển vi ánh sáng.
Các chức năng của viên nang và chất nhờn được đưa ra dưới đây:
1. Nó ngăn ngừa làm khô các tế bào, bằng cách liên kết các phân tử nước.
2. Nó ngăn chặn sự gắn kết của vi khuẩn (virus, tấn công vi khuẩn) vào các tế bào vi khuẩn.
3. Nó cung cấp độc lực (khả năng tạo bệnh) bằng cách ức chế sự nhận biết của chúng và sự phá hủy sau đó của thực bào (tiểu thể bạch cầu hoặc WBC).
4. Nó giúp vi khuẩn gây bệnh bám vào các thụ thể trên bề mặt mô chủ bị nhiễm bệnh.
5. Nó cung cấp sự tuân thủ giữa các tế bào vi khuẩn khác nhau.
6. Vỏ bọc:
Vỏ bọc là các cấu trúc hình ống rỗng dài, bao gồm các chuỗi tế bào vi khuẩn hình thành 'trichomes' (Hình 2.9). Vi khuẩn có vỏ bọc phổ biến trong môi trường nước ngọt rất giàu chất hữu cơ, chẳng hạn như dòng nước ô nhiễm, bộ lọc nhỏ giọt và chất phân hủy bùn hoạt tính trong các nhà máy xử lý nước thải. Vỏ bọc có thể dễ dàng hình dung, khi một số tế bào, được gọi là 'tế bào swarmer được đánh dấu' của bộ ba di chuyển ra khỏi vỏ để lại vỏ bọc trống.

7. Ưu-thecae:
Ưu điểm là các phần mở rộng tế bào chất từ tế bào vi khuẩn, hình thành các phần phụ như sợi nấm, cuống hoặc chồi. Chúng có đường kính nhỏ hơn các tế bào trưởng thành, chứa tế bào chất và được giới hạn bởi thành tế bào. Vi khuẩn có pros-thecae được gọi là vi khuẩn tuyến tiền liệt.
Không giống như sự phân chia tế bào ở các vi khuẩn khác, diễn ra thông qua quá trình phân hạch nhị phân tạo ra hai tế bào vi khuẩn con tương đương mà ở tuyến tiền liệt liên quan đến việc hình thành một tế bào con mới với tế bào mẹ vẫn giữ được bản sắc sau khi quá trình phân chia tế bào hoàn tất.
"Hyphae" là phần phụ của tế bào được hình thành từ sự mở rộng tế bào trực tiếp của tế bào mẹ có chứa thành tế bào, màng tế bào, ribosome và đôi khi là DNA. Nó được hình thành bởi sự kéo dài của một sự phát triển nhỏ của tế bào vi khuẩn tại một điểm duy nhất.
Sự tăng trưởng như vậy tại một điểm được gọi là "tăng trưởng cực" trái ngược với "sự tăng trưởng xen kẽ" diễn ra ở tất cả các vi khuẩn khác, trong đó sự phát triển của vi khuẩn diễn ra trên khắp bề mặt. Vào cuối của sợi nấm, một chồi được hình thành, nó mở rộng và tạo ra một lá cờ.
DNA trong tế bào mẹ sao chép và một bản sao DNA tròn được di chuyển xuống theo chiều dài của sợi nấm vào chồi. Một vách ngăn chéo sau đó hình thành tách chồi vẫn đang phát triển từ tế bào mẹ và tế bào mẹ.
Bây giờ, chồi vỡ ra khỏi tế bào mẹ và bơi đi như một tế bào con. Sau đó, tế bào con mất đi tế bào của nó và dần dần phát triển để trở thành tế bào mẹ sẵn sàng cho sự hình thành của sợi nấm và chồi.
"Thân cây" là phần phụ của tế bào được sử dụng bởi một số vi khuẩn thủy sinh để gắn vào chất nền rắn. Ở phần cuối của thân cây có một cấu trúc gọi là 'giữ vững', theo đó thân cây được gắn vào đế. Bên cạnh việc cung cấp sự gắn kết, thân cây cũng làm tăng diện tích bề mặt của tế bào vi khuẩn, giúp nó hấp thụ chất dinh dưỡng và loại bỏ chất thải.
Ở một số vi khuẩn, thân của một số cá thể vẫn được gắn vào để tạo thành 'hoa hồng'. Sự phân chia tế bào ở vi khuẩn rình rập xảy ra do sự kéo dài của tế bào theo hướng ngược lại với hướng của thân cây sau đó là sự phân hạch. Một cột cờ đơn được hình thành ở cực đối diện với thân cây.
Các tế bào con được gắn cờ được hình thành như vậy được gọi là "swarmer". Nó tách ra khỏi tế bào mẹ không được gắn cờ, bơi xung quanh và gắn vào một bề mặt mới tạo thành một cuống mới ở cực được gắn cờ. Flagellum sau đó bị mất.
"Nụ" là phần phụ của tế bào, chúng trực tiếp phát triển thành tế bào con từ tế bào mẹ mà không có sự hình thành của sợi nấm.
8. Thành tế bào:
Thành tế bào là cấu trúc cứng nhắc ngay lập tức bao quanh màng tế bào của tế bào vi khuẩn. Màng tế bào của tế bào vi khuẩn là một lớp kép phospholipid tương tự như tất cả các tế bào động vật và thực vật khác (Hình 2.11).

Trong khi các tế bào động vật thiếu thành tế bào, tế bào thực vật và tế bào vi khuẩn có thành tế bào bao quanh màng tế bào. Tuy nhiên, không giống như thành tế bào của tế bào thực vật, có thành phần chủ yếu là cellulose, thành tế bào của tế bào vi khuẩn được cấu tạo chủ yếu từ các lớp peptidoglycan (Hình 2.10).

Trước đó, peptidoglycan cũng được gọi là murein hoặc mucopeptide. Peptidoglycan bao gồm chủ yếu là axit amin và đường (peptido: chuỗi axit amin; glycan: đường). Hai loại đường là N-acetylglucosamine (NAGA) và axit N-acetylmuramic (NAMA) được liên kết với nhau thông qua các liên kết glycosid (liên kết β-1-4) để tạo thành chuỗi đường song song dài.
Đi kèm với các phân tử NAMA là các chuỗi bên ngắn gồm bốn hoặc năm axit amin liên kết với nhau thông qua các liên kết peptide. Thông thường các axit amin là alanine, axit glutamic và axit lysine hoặc diaminopimelic (DAP). Các peptide ngắn này tạo thành các liên kết chéo giữa các chuỗi đường liền kề để tạo thành một mạng hoặc lớp.
Mặc dù các liên kết glycosid trong chuỗi đường rất mạnh, nhưng những thứ này không thể cung cấp độ cứng theo mọi hướng. Độ cứng cao thu được, chỉ khi các chuỗi này được liên kết chéo bởi các liên kết peptide trong các peptide ngắn liên kết ngang.
Thành phần của thành tế bào không giống nhau đối với tất cả các vi khuẩn; thay vào đó là sự khác biệt đáng chú ý trong thành phần của nó, dựa trên đó vi khuẩn đã được nhóm lại như được đưa ra dưới đây:
A. Dựa trên nhuộm Gram:
Dựa trên sự khác biệt về thành phần của thành tế bào, một kỹ thuật nhuộm đặc biệt gọi là "nhuộm gram", được đặt theo tên của Tiến sĩ Christian Gram, người đã phát triển nó, phân biệt vi khuẩn thành hai nhóm như sau:
(1) Vi khuẩn gram dương:
Sau khi nhuộm vi khuẩn bằng pha lê tím, nếu thành tế bào của nó chống lại sự mất màu bằng cách rửa bằng chất khử màu (ethanol hoặc acetone), đó là vi khuẩn gram dương. Ví dụ: Bacillus, Staphylococcus.
(2) Vi khuẩn gram âm:
Sau khi nhuộm vi khuẩn bằng pha lê tím, nếu thành tế bào của nó cho phép trải qua quá trình khử màu bằng cách rửa bằng chất khử màu (ethanol hoặc acetone), đó là vi khuẩn gram âm. Ví dụ: Escherichia, Salmonella, Vibrio.
Phân biệt vi khuẩn thành các nhóm gram dương và gram âm cung cấp manh mối quan trọng nhất để tiến xa hơn theo hướng thích hợp để xác định vi khuẩn chưa biết.
Thành tế bào gram dương:
Thành tế bào gram dương rất đơn giản. Nó chủ yếu được làm bằng một lớp peptidoglycan dày, chiếm khoảng 90% của thành tế bào. Lớp dày này làm cho vi khuẩn gram dương chống lại sự thẩm thấu của thẩm thấu.
Thành này cũng có thể chứa axit teichoic, axit lipoteichoic và protein M, là những kháng nguyên bề mặt chính của thành tế bào gram dương. Axit teichoic tích điện âm và chịu trách nhiệm một phần cho điện tích âm của bề mặt tế bào của vi khuẩn. Nó cũng giúp trong việc đi qua các ion qua thành tế bào.
Axit lipoteichoic là axit teichoic, liên kết với lipit màng. Protein M giúp một số vi khuẩn gây bệnh bằng cách ngăn chặn sự nhấn chìm của chúng bằng WBC. Các kháng sinh, penicillin (được sản xuất bởi nấm, Penicillium notatum) và cephalosporin tiêu diệt vi khuẩn gram dương bằng cách ngăn chặn sự tổng hợp peptidoglycan. Lysozyme (một loại enzyme có trong nước mắt, nước bọt, albumin trứng và chất nhờn của cá) giết chết vi khuẩn gram dương bằng cách phá vỡ liên kết glycosid (liên kết-1-4) giữa NAGA và NAMA.
Thành tế bào gram âm:
Thành tế bào gram âm rất phức tạp và dễ vỡ hơn thành tế bào gram dương. Nó có một lớp peptidoglycan mỏng, chỉ chiếm khoảng 10% của thành tế bào. Nó được bao quanh bởi một lớp bảo vệ dày gọi là lớp lipopolysacarit (LPS), chống lại sự xâm nhập của một số hóa chất độc hại tiềm tàng vào tế bào vi khuẩn.
Lớp LPS bao gồm 1) lớp lipoprotein và 2) lớp màng ngoài. Một bề mặt của lớp lipoprotein được gắn trực tiếp vào lớp peptidoglycan và bề mặt còn lại kéo dài vào màng ngoài.
Nó có chức năng như một mỏ neo giữa lớp peptidoglycan và màng ngoài. Màng ngoài là màng kép phospholipid nhúng protein với cấu trúc 'màng đơn vị' tương tự màng tế bào. Lá trong của màng ngoài là một lớp các phân tử phospholipid đơn thuần.
Tuy nhiên, không giống như màng đơn vị, một số phân tử phospholipid ở lá ngoài được thay thế bằng các phân tử LPS. Mỗi phân tử LPS được tạo thành từ một phần lipid gọi là 'lipid A' liên kết cộng hóa trị với một polysacarit giống như tóc.
Khi vi khuẩn gây bệnh gram âm bị tiêu diệt bên trong cơ thể vật chủ, lipid A, một thành phần bên trong của vi khuẩn gram âm, được giải phóng, gợi ra các phản ứng độc hại và các triệu chứng bệnh. Đó là lý do tại sao, lipid A còn được gọi là 'endotoxin' (endo: Internal).
Ngược lại, 'exotoxin' là chất độc hại được tổng hợp bên trong tế bào vi khuẩn và thải ra bên ngoài, gây bệnh, nếu có hoặc xâm nhập vào cơ thể vật chủ. Không giống như các lipit bình thường khác như triglyceride, được tạo thành từ ba axit béo liên kết với glycerol, lipid A được tạo thành từ các axit béo liên kết với một disacarit gồm hai photphat NAGA.
Các axit béo trong lipid A thường là axit caproic, lauric, myristic, palmitic và stearic. Các polysacarit, được liên kết với lipid A ở đầu kia của disacarit, được làm bằng hai phần; 1) polysacarit lõi bên trong và 2) O-polysacarit bên ngoài.
Màng ngoài cũng chứa các protein gọi là 'porins', chứa ba tiểu đơn vị giống hệt nhau. Mỗi tiểu đơn vị có dạng hình trụ với lỗ rỗng trung tâm, qua đó vật liệu có thể đi qua. Vì vậy, các porin làm cho màng ngoài tương đối thấm.
Hiên có hai loại; 1) các porin không đặc hiệu, tạo thành các kênh chứa đầy nước cho các chất nhỏ thuộc bất kỳ loại nào đi qua và 2) các porin cụ thể, có các vị trí liên kết cụ thể cho các chất cụ thể đi qua.
Một số vật liệu (dung dịch, ống tiêm, kim tiêm, v.v.), mặc dù không có vi khuẩn gây bệnh bằng cách khử trùng bằng nhiệt, vẫn giữ lại các mảnh vụn còn sót lại của thành tế bào của vi khuẩn gram âm được tạo ra bằng cách tiêu diệt chúng. Nếu được đưa vào cơ thể vật chủ, mảnh vỡ thành tế bào này có thể gây ra các triệu chứng bệnh. Mảnh vỡ thành tế bào bị vỡ này, có thể gây ra các triệu chứng bệnh, được gọi là 'pyrogen'.
B. Dựa trên nhuộm nhanh axit:
Dựa trên sự khác biệt về thành phần của thành tế bào, một kỹ thuật nhuộm đặc biệt khác gọi là "nhuộm nhanh axit" phân biệt vi khuẩn thành hai nhóm như sau:
(1) Vi khuẩn nhanh axit:
Một loại vi khuẩn, cực kỳ khó nhuộm, nhưng một khi đã nhuộm màu, thì việc loại bỏ vết bẩn ra khỏi tế bào cũng khó khăn không kém, ngay cả khi sử dụng mạnh mẽ axit-rượu làm chất khử màu, là một loại vi khuẩn có tính axit nhanh vi khuẩn).
Ví dụ: Mycobacterium spp. [M bệnh lao (vi khuẩn lao), M. leprae (vi khuẩn bệnh phong), M. smegmatis (vi khuẩn tự nhiên của smegma), M. marinum (vi khuẩn lao của cá biển)]. Chúng sở hữu một thành tế bào sáp dày làm từ vật liệu lipid.
(2) Vi khuẩn không axit nhanh:
Một loại vi khuẩn, dễ bị nhuộm màu và cũng dễ bị phân hủy bởi rượu axit là chất khử màu, là một loại vi khuẩn không axit nhanh (vi khuẩn không ưa axit). Ví dụ: Tất cả các vi khuẩn khác trừ Mycobacterium spp. Ở những vi khuẩn này, thành tế bào không có sáp dày.
Phân biệt vi khuẩn thành các nhóm axit nhanh và không axit nhanh cung cấp manh mối quan trọng nhất để xác định Mycobacterium spp.
Biến thể thiếu tường:
Một số vi khuẩn tự nhiên tồn tại mà không có thành tế bào. Mycoplasma, là vi khuẩn nhỏ nhất, không có thành tế bào. Nó được bảo vệ khỏi sự thẩm thấu của thẩm thấu bởi sự hiện diện của sterol trong màng tế bào và bằng cách áp dụng sự tồn tại của ký sinh trùng trong môi trường thuận lợi về mặt thẩm thấu của vật chủ nhân chuẩn.
Tương tự, một số vi khuẩn biển thiếu thành tế bào, trong đó sự phân giải thẩm thấu bị ngăn chặn bởi nồng độ muối cao của nước biển. Hầu hết các vi khuẩn gram dương mất thành tế bào của chúng (peptidoglycan) với sự hiện diện của lysozyme.
Tuy nhiên, các tế bào của chúng tồn tại nếu chúng có mặt trong môi trường đẳng trương như vết thương có mủ. Những vi khuẩn thiếu tường này chỉ bao gồm các nguyên sinh chất được bao bọc bởi màng tế bào được gọi là 'protoplast' hoặc 'L-Forms' (L cho Viện Lister ở London).
Khi lysozyme được loại bỏ, chúng có thể tiếp tục tăng trưởng bằng cách tổng hợp thành tế bào của chúng. Nếu protoplast giữ lại các mảnh của thành tế bào, sao cho tế bào có hình dạng amip, nhô ra ở nơi không có thành tế bào, nó được gọi là 'spheroplast'.
Spheroplast cũng được hình thành, khi peptidoglycan của thành tế bào gram âm được loại bỏ bằng cách xử lý với lysozyme. Ở đây, nguyên sinh chất được bao bọc bởi hai lớp, màng tế bào và lớp LPS.
Chức năng của vách tế bào:
(a) Mẫu bảo vệ Lysis:
Nó bảo vệ các tế bào vi khuẩn khỏi ly giải thẩm thấu, vì hầu hết chúng vẫn ở trong môi trường hypotonic. Điều này là bởi vì; nồng độ các chất hòa tan bên trong tế bào vi khuẩn cao hơn đáng kể so với trong môi trường. Điều này phát triển một áp lực đáng kể bên trong các tế bào, có thể tương đương với áp suất bên trong lốp ô tô. Thành tế bào giúp chịu được áp lực này.
(b) Hình dạng và độ cứng:
Nó cung cấp hình dạng và độ cứng cho các tế bào vi khuẩn.
9. Không gian Periplasmic:
Đó là khoảng trống mỏng giữa thành tế bào và màng tế bào (Hình 2.10 và 2.11). Nó chứa đầy huyết tương gọi là periplasm. Các periplasm chứa một số protein, trong đó ba là quan trọng.
Chúng là, 1) enzyme thủy phân, giúp phân hủy ban đầu các phân tử thức ăn của vi khuẩn, 2) protein liên kết, bắt đầu quá trình vận chuyển cơ chất và 3) chemoreceptor, có liên quan đến phản ứng hóa trị của vi khuẩn. Hóa trị có nghĩa là di chuyển tới hoặc ra khỏi một hóa chất.
10. Màng tế bào:
Đó là một màng mỏng bên dưới thành tế bào bao bọc hoàn toàn nguyên sinh chất. Nó cũng được gọi là "màng tế bào chất" hay "màng plasma". Nó là một lớp kép phospholipid nhúng protein (Hình 2.11).
Hai loại protein được nhúng trong hai lớp. Một loại protein được giữ vững chắc trong hai lớp và được gọi là "protein tách rời". Các loại protein khác được neo lỏng lẻo trên bề mặt của lớp kép và được gọi là 'protein ngoại vi'.
Các thành phần của màng không tĩnh; đúng hơn là họ năng động và luôn trong trạng thái chuyển động như một 'chất lỏng'. Các protein cung cấp cho màng có hình dạng 'khảm'.
Đó là lý do tại sao; màng đơn vị thường được thể hiện thông qua "mô hình khảm chất lỏng". Bộ đôi phospholipids bao gồm hai lớp đối diện nhau, mỗi lớp bao gồm một lớp phân tử phospholipids. Mỗi phân tử phospholipids có một đầu và một cặp đuôi nổi lên từ nó.
Đầu được tích điện và ưa nước (ưa nước), mà nó vẫn hướng tới môi trường nước. Các đuôi là không tích điện và kỵ nước (ghét nước), mà chúng vẫn ẩn trong màng cách xa môi trường nước. Một màng như vậy được gọi là "màng đơn vị".
Sterol, hiện diện trong màng đơn vị của tất cả các sinh vật nhân chuẩn là tác nhân tăng cường, không có trong màng đơn vị của hầu như tất cả các vi khuẩn. Tuy nhiên, các phân tử tương tự như sterol, được gọi là 'humanoids' có trong một số vi khuẩn, có vai trò tương tự như các sterol trong sinh vật nhân chuẩn. Một trong những hình người quan trọng là 'Diploptene', có 30 nguyên tử carbon.
Chức năng của màng tế bào:
(a) Rào cản thấm:
Phần bên trong của tế bào (tế bào chất) bao gồm dung dịch muối, đường, axit amin, vitamin, coenzyme và nhiều loại vật liệu hòa tan khác. Bản chất kỵ nước của màng tế bào làm cho nó & hàng rào chặt chẽ.
Mặc dù một số phân tử kỵ nước nhỏ có thể đi qua màng bằng cách khuếch tán, các phân tử ưa nước và tích điện (hầu hết các phân tử hòa tan trong nước) không dễ dàng đi qua nó. Các phân tử này được vận chuyển qua nó bởi các hệ thống vận chuyển cụ thể.
Ngay cả một chất nhỏ như ion hydro (H + ) cũng không khuếch tán qua màng tế bào. Phân tử duy nhất thâm nhập vào màng là nước, đủ nhỏ và không tích điện để đi qua giữa các phân tử phospholipids.
Do đó, màng tế bào là một rào cản quan trọng ngăn cách bên trong tế bào với môi trường của nó và ngăn chặn sự rò rỉ thụ động của các thành phần tế bào chất vào hoặc ra khỏi tế bào. Nếu màng bị phá vỡ, tính toàn vẹn của tế bào bị phá hủy, nội dung bên trong bị rò rỉ ra môi trường và tế bào chết.
(b) Rào cản chọn lọc:
Màng tế bào cũng là một rào cản chọn lọc cao. Các protein cụ thể, được gọi là "hệ thống vận chuyển màng" có mặt trên và trong màng tế bào cho phép và tạo điều kiện vận chuyển các chất cụ thể qua nó bằng cách vận chuyển chủ động và thụ động.
Về mặt cấu trúc, các hệ thống vận chuyển màng là các protein được tạo thành từ 12 vòng xoắn alpha cuộn qua lại qua màng để tạo thành một kênh, qua đó chất được vận chuyển được đưa vào hoặc ra khỏi tế bào.
Những protein này có thể liên kết với các chất cụ thể sẽ được vận chuyển và di chuyển chúng vào hoặc ra khỏi tế bào qua màng theo yêu cầu. Điều này cho phép một tế bào vi khuẩn tập trung các chất cụ thể bên trong tế bào và loại bỏ các chất cụ thể từ tế bào ra bên ngoài.
Có ba loại protein vận chuyển màng, chẳng hạn như 1) Uniporters, chỉ vận chuyển một loại phân tử theo một hướng qua màng, 2) Symporter, vận chuyển một loại phân tử và luôn vận chuyển một loại phân tử khác cùng với nó cùng hướng và 3) Antiporters, vận chuyển một loại phân tử và luôn vận chuyển một loại phân tử khác theo hướng ngược lại.
Việc vận chuyển các chất có thể diễn ra theo ba cách, chẳng hạn như 1) Vận chuyển đơn giản, trong đó việc vận chuyển được thực hiện qua màng bằng năng lượng từ động lực proton, 2) Dịch chuyển nhóm, trong đó chất vận chuyển bị biến đổi hóa học trong quá trình đi qua xuyên qua màng và 3) Vận chuyển ABC (băng liên kết ATP), trong đó protein liên kết periplasmic trong periplasm liên kết đặc biệt với chất, protein bao màng, kéo dài qua màng vận chuyển qua màng và tế bào chất Protein thủy phân ATP có trong tế bào chất cung cấp năng lượng cho sự kiện vận chuyển.
(c) Thủy phân ngoại bào:
Màng tế bào tiết ra nhiều enzyme ra bên ngoài để phân hủy ngoại bào của các hạt thức ăn lớn thành các phân tử, có thể đi vào tế bào qua màng.
(d) Hô hấp:
Vi khuẩn không có ty thể. Do đó, hô hấp tế bào, để lấy năng lượng từ nguyên liệu thực phẩm, được thực hiện trong màng tế bào, nơi chứa các enzyme của chuỗi hô hấp. Nó là một thiết bị để bảo tồn năng lượng trong tế bào.
Màng có thể tồn tại ở dạng 'tích điện', trong đó sự phân tách các proton (H + ) từ các ion hydroxyl (OH-) xảy ra trên bề mặt của nó. Sự phân tách điện tích này là một dạng năng lượng trao đổi chất, tương tự như năng lượng tiềm năng có trong pin sạc.
Trạng thái năng lượng của màng, được gọi là 'lực thúc đẩy proton' (PMF), chịu trách nhiệm điều khiển nhiều chức năng cần năng lượng trong tế bào, bao gồm một số dạng vận chuyển, vận động và sinh tổng hợp tiền tệ năng lượng của tế bào, ATP .
(e) Quang hợp:
Vi khuẩn quang điện thực hiện quá trình quang hợp bởi các bào quan đặc biệt gọi là "sắc tố" được hình thành bởi trí tưởng tượng của màng tế bào. Những bào quan này chứa enzyme và sắc tố để quang hợp.
(f) Hấp thụ các chất dinh dưỡng:
Các chất dinh dưỡng xung quanh được hấp thụ trực tiếp vào các tế bào vi khuẩn thông qua màng tế bào. "Mesosome" là sự xâm lấn màng tế bào của màng tế bào vào tế bào chất, làm tăng diện tích bề mặt để hấp thu hiệu quả.
(g) Sinh sản:
Vi khuẩn sinh sản bằng phân hạch nhị phân. Mỗi tế bào phân chia thành hai tế bào con. Trong quá trình này, màng tế bào tạo thành thành tế bào mới giữa các tế bào con đang phân chia.
11. Tế bào chất:
Chất giống như chất lỏng nhớt, lấp đầy khoang bên trong màng tế bào, được gọi là tế bào chất (Hình 2.10 và 2.11). Nó là một chất lỏng làm từ nước, enzyme và các phân tử nhỏ. Bị đình chỉ trong chất lỏng là một vùng tương đối trong suốt của vật liệu nhiễm sắc thể gọi là 'nucleoid', hạt ribosome và các thể vùi tế bào chất, lưu trữ năng lượng.
12. Hạt nhân:
Chính khối lượng DNA tổng hợp tạo nên nhiễm sắc thể của vi khuẩn (Hình 2.12). Vì vi khuẩn là sinh vật nhân sơ, DNA của chúng không bị giới hạn trong nhân gắn màng, giống như ở sinh vật nhân chuẩn.

Tuy nhiên, nó không nằm rải rác trong tế bào, mà nó có xu hướng tập hợp thành một cấu trúc riêng biệt trong tế bào vi khuẩn và được gọi là "nucleoid" (Hình 2.12). Ribosome không có trong khu vực của nucleoid, bởi vì DNA nucleoid tồn tại ở dạng giống như gel loại trừ vật chất hạt.
Hầu hết các vi khuẩn chỉ chứa một nhiễm sắc thể duy nhất, bao gồm một chuỗi DNA kép đơn, mặc dù một số vi khuẩn có DNA nhiễm sắc thể tuyến tính. Thông thường, DNA nhiễm sắc thể được xoắn và gấp lại thành 'DNA siêu cuộn', để chứa nó trong không gian nhỏ của nucleoid.
DNA siêu cuộn nhỏ gọn hơn DNA tuần hoàn tự do. Vi khuẩn chỉ chứa một bản sao duy nhất của mỗi gen trên nhiễm sắc thể của chúng và do đó là "đơn bội" về mặt di truyền. Ngược lại, sinh vật nhân chuẩn chứa hai bản sao của mỗi gen trong nhiễm sắc thể của chúng và do đó là lưỡng bội di truyền.
Gene có thể được định nghĩa là bất kỳ phân đoạn nào của DNA, chịu trách nhiệm cho một đặc tính cụ thể của sinh vật chủ. Mỗi nhân vật của một sinh vật được chỉ định bởi một gen cụ thể, bao gồm một đoạn DNA cụ thể.
DNA nhiễm sắc thể của vi khuẩn chứa 'gen giữ nhà' cần thiết, cần thiết cho sự sống sót cơ bản của vi khuẩn. DNA này được phân phối giữa hai tế bào con trong quá trình phân chia tế bào.
Bên cạnh các gen nhiễm sắc thể, còn có các gen không nhiễm sắc thể có mặt bên ngoài nhiễm sắc thể. Ví dụ, các plasmid chứa các gen cụ thể, tạo ra các đặc tính đặc biệt cho vi khuẩn. Tổng số tất cả các gen có trong một tế bào được gọi là "bộ gen" của nó.
13. Plasmid:
Đây là một số ít DNA không nhiễm sắc thể được tìm thấy trong tế bào chất của một số vi khuẩn (Hình 2.12). Mỗi plasmid bao gồm một đoạn DNA sợi đôi tròn nhỏ, có thể sao chép độc lập với nhiễm sắc thể. Nhiều plasmid khá nhỏ, nhưng một số khá lớn.
Tuy nhiên, không có plasmid nào lớn như nhiễm sắc thể. Plasmid không có gen cần thiết cho sự phát triển hoặc sống sót của vi khuẩn. Họ có thể có các gen chịu trách nhiệm kháng kháng sinh, sản xuất độc tố hoặc sản xuất các phần phụ trên bề mặt cần thiết cho sự gắn kết và thiết lập nhiễm trùng.
Do đó, các plasmid chứa các gen mang tính chất đặc biệt cho vi khuẩn, không giống như DNA nhiễm sắc thể, chứa các gen giữ nhà thiết yếu cần thiết cho sự sống sót cơ bản của vi khuẩn.
Một số plasmid, có thể tích hợp vào DNA nhiễm sắc thể chủ được gọi là 'episomes'. Plasmid cũng có thể được loại bỏ khỏi tế bào vi khuẩn chủ bằng quá trình gọi là 'chữa bệnh'. Một số plasmid được sử dụng rộng rãi trong thao tác gen và kỹ thuật di truyền.
14. Ribosome:
Đây là những cơ thể dạng hạt có trong tế bào chất, trong đó quá trình tổng hợp protein diễn ra (Hình 2.3). Chúng rất nhiều đến nỗi, sự hiện diện của chúng mang lại cho tế bào chất một vẻ ngoài tối tăm. Kích thước của mỗi ribosome vi khuẩn là 70 S (70 Svedberg), bao gồm hai tiểu đơn vị kích thước 50S và 30S. Ngược lại, kích thước của mỗi ribosome nhân chuẩn là 80 S bao gồm hai tiểu đơn vị kích thước 60S và 40S.
15. Vắc xin khí:
Không bào khí là các cấu trúc chứa đầy khí hình trục chính được tìm thấy trong tế bào chất (Hình 2.3 và 2.13). Chúng là những ống hình trụ rỗng, được đóng bởi một nắp hình nón rỗng ở mỗi đầu. Số lượng của chúng trong một tế bào có thể thay đổi từ vài đến hàng trăm.

Their length and width vary in different bacteria, but the vacuoles of any given bacteria are more or less of constant size. They are hollow, but rigid. They are impermeable to water and solutes, but are permeable to most gases. Therefore, the composition of the gas inside them is the same as that of the gas, in which the organism is suspended.
Gas is present in the vacuoles at about 1 atm. sức ép. Therefore, rigidity of gas vacuole membrane is essential for the vacuole to resist the pressure exerted on it from outside. Due to this, it is composed of proteins able to form a rigid membrane rather than of lipids, which would form a fluid, highly mobile membrane.
There are two types of proteins, which make the membrane. They are 1) GvpA and 2) GvpC (Gas vacuole protein A and C). GvpA is the major gas vacuoles protein, which is small and highly hydrophobic (Figure 2.13). It is the shell protein and makes up 97% of the total protein of the gas vacuoles.
Gas vacuoles are made of several GvpA protein molecules aligned as parallel 'ribs' forming a watertight surface. GvpA proteins fold as a (3-sheet and thus, give considerable rigidity to the overall vacuole structure. GvpC is a larger protein, but is present in much smaller amount.
It strengthens the shell of the gas vacuoles by acting as cross-linker, binding several GvpA ribs together like a clamp. The final shape of the gas vacuoles, which can vary in different organisms from long and thin to short and fat is a function of how the GvpA and GvpC proteins are arranged to form the intact vacuoles.
As the gas vacuoles are filled with gases, their density is about 5- 20% of that of the cell proper. Thus, the intact gas vacuoles decrease the density of the bacteria cells, thereby conferring buoyancy to them.
Most of the floating aquatic bacteria have gas vacuoles, which confer buoyancy and help them to float on the surface of water or anywhere in the water column. Gas vacuoles also act as a means of motility, allowing cells to float up and down in the water column in response to environmental factors.
For example, aquatic phototrophic organisms benefit from gas vacuoles, which help them to adjust their position rapidly in the water column to regions where light intensity for photosynthesis is optimal. The gas vacuole membrane cannot resist high hydrostatic pressure and can collapse, leading to a loss of buoyancy. Once collapsed, gas vacuoles cannot be reinflated.
16. Inclusions:
The non-living reserve materials suspended in the cytoplasm are called 'inclusions' (Figure 2.3). Most of the inclusions are bounded by a thin non-unit membrane consisting of lipids separating them from the surrounding cytoplasm.
Their function is the storage of energy or structural building blocks. Different bacteria contain different types of inclusions.
Some of the common inclusions found in bacteria cells are described below:
(a) Poly-β-hydroxybutyric Acid (PHB) Granules:
These are granules consisting of poly- β-hydroxybutyric acid (PHB), which is a lipid-like substance. Monomers of β-hydroxybutyric acid are bonded to each other in a chain by ester linkages to form the polymer of poly- β-hydroxybutyric acid. The polymers aggregate into granules.
(b) Poly-β-hydroxyalkanoate (PHA) Granules:
It is a collective term used to describe all classes of carbon/energy storage polymers found in bacterial cytoplasm. The most common PHA granules are the PHB granules. The length of the monomers in a polymer can vary, from as short as C 4 (in PHB granules) to as long as C 18 (in other granules) in many bacteria.
(c) Volutin Granules:
These are granules composed of polyphosphates. The basic dye, toluidine blue becomes reddish violet, when it combines with the polyphosphate granules. This phenomenon of color change is called 'metachromasy'. Therefore, volutin granules are also called 'metachromatic granules'.
(d) Glycogen Granules:
These are starch-like polymers made of glucose subunits. Glycogen granules are usually smaller than PHB granules. These are storage depots of carbon and energy in bacteria cells.
(e) Sulphur Granules:
These are granules of elemental sulphur produced by its accumulation in the cytoplasm. Elemental sulphur comes from the oxidation of reduced sulphur compounds like hydrogen sulphide and thiosulphate. The granules remain as long as the source of reduced sulphur is present. As the source disappears, being utilized by the bacteria cells, the granules also disappear slowly.
(f) Magnetosomes:
Magnetosomes are granules of the iron mineral magnetite, Fe 3 O 4 . They impart a permanent magnetic dipole to a bacteria cell, allowing it to respond to magnetic fields. A membrane containing phospholipids, proteins and glycoproteins surround each magnetosome.
The membrane proteins play a role in precipitating Fe 3+ (brought into the cell in soluble form by chelating agents) as Fe 3 0 4 in the developing magnetosomes. The morphology of magnetosomes appears to be species-specific, varying in shape from square to rectangular to spike-shaped in certain bacteria.
Magnetosomes are primarily found in aquatic bacteria. Bacteria that produce magnetosomes orient and migrate along geomagnetic field. This phenomenon is called 'magnetotaxis'. Bacteria, which show magnetotaxis, are called 'magnetotactic bacteria'.
There is no evidence chat magnetotactic bacteria employ the sensory systems of chemotactic or phototactic bacteria. Instead, the alignment of magnetosomes in a bacteria cell simply imparts magnetic properties to it, which then orients the cell in a particular direction in its environment. Thus, a better term to describe these organisms is ' magnetic bacteria'.
17. Endospore:
It is a differentiated cell formed within the cell of certain gram-positive rod-shaped bacteria, which can withstand severe adverse conditions like heat, cold, radiation, ageing, toxic chemicals and lack of nutrients.
Mostly, the following three genera of rod-shaped bacteria can produce endospores in adverse conditions:
I. Aerobic rod-shaped bacteria
1. Bacillus spp.
II. Anaerobic rod-shaped bacteria
2. Clostridium spp.
3. Desulfotomaculum spp.
Structure of Endospore:
The structure of the endospore is much more complex than that of the vegetative cell in that it has many layers. The layers from outside towards inside are given below (Figure 2.14).

(a) Exosporium:
It is the outermost layer, which is a thin, delicate covering, made of proteins.
(b) Spore coat:
It is made of layers of spore-specific proteins. It acts like a sieve that excludes large toxic molecules like lysozyme, is resistant to many toxic molecules and may also contain enzymes that are involved in the germination of the spore.
(c) Cortex:
It is made of loosely cross-linked peptidoglycan.
(d) Core:
It contains the usual structures of bacteria cell, which includes the cell wall (core wall), enclosing the spore protoplast. The spore protoplast consists of the cell membrane, cytoplasm, nucleoid, ribosomes, inclusions and so on.
Thus, the spore differs structurally from the vegetative cell primarily in the kinds of structures found outside the core wall.
Thuộc tính của lõi Endospore:
Cốt lõi của một endospore trưởng thành khác rất nhiều với tế bào sinh dưỡng, từ đó nó được hình thành.
Các thuộc tính của lõi endospore như sau:
(a) Một chất hóa học đặc trưng của endospores, nhưng không có trong tế bào sinh dưỡng là axit dipicolinic (DPA). Nó nằm trong lõi.
(b) Các bào tử cũng chứa nhiều ion canxi, hầu hết được kết hợp với axit dipicolinic. Phức hợp axit canxi-dipicolinic của lõi chiếm khoảng 10% trọng lượng khô của endospore.
(c) Bên cạnh việc có hàm lượng axit canxi-dipicolinic (Ca-DPA) dồi dào, lõi ở trạng thái mất nước một phần. Lõi của một endospore trưởng thành chỉ chứa 10-30% hàm lượng nước của tế bào sinh dưỡng và do đó, tính nhất quán của tế bào chất lõi là của gel.
Mất nước làm tăng đáng kể khả năng chịu nhiệt của endospore cũng như chống lại các hóa chất, chẳng hạn như hydro peroxide (H 2 O 2 ) và làm cho các enzyme có trong lõi không hoạt động.
(d) Ngoài hàm lượng nước thấp của bào tử, độ pH của tế bào chất lõi thấp hơn khoảng một đơn vị so với tế bào sinh dưỡng.
(e) Lõi chứa hàm lượng protein đặc hiệu lõi cao gọi là 'protein bào tử hòa tan axit nhỏ' (SASPs). Chúng được tổng hợp trong quá trình đầu cơ và có hai chức năng. Đầu tiên, chúng liên kết chặt chẽ với DNA trong lõi và bảo vệ nó khỏi thiệt hại tiềm tàng từ bức xạ UV, hút ẩm và nhiệt khô. Thứ hai, chúng hoạt động như nguồn carbon và năng lượng cho sự phát triển của tế bào sinh dưỡng mới bằng quá trình nảy mầm.
Chu kỳ bào tử:
Trong điều kiện bất lợi, một bào tử được tạo ra trong một tế bào sinh dưỡng do mất nước và co lại của nội dung tế bào (Hình 2.14). Bào tử này, được hình thành bên trong tế bào vi khuẩn, được gọi là 'endospore'. Nếu các điều kiện bất lợi trở nên tồi tệ hơn, các tế bào sẽ phá vỡ giải phóng nội nhũ, giờ đây trở thành một tế bào không hoạt động độc lập gọi là 'bào tử'.
Quá trình, theo đó một tế bào sinh dưỡng tạo ra một bào tử, được gọi là "sinh bào tử" hay "bào tử". Các bào tử vẫn ở dạng không hoạt động cho đến khi các điều kiện bất lợi vẫn tồn tại. Khi trở lại điều kiện thuận lợi, bào tử trở lại dạng thực vật và hoạt động trở lại.
Các lớp bao phủ vỡ bào tử và bào tử tạo ra một tế bào sinh dưỡng hoạt động trao đổi chất. Quá trình, qua đó một bào tử thay đổi thành một tế bào sinh dưỡng, được gọi là "nảy mầm".
Quá trình thay đổi theo chu kỳ từ dạng thực vật sang bào tử trong điều kiện bất lợi và trở lại dạng thực vật trong điều kiện thuận lợi được gọi là "chu kỳ bào tử".
Hai nửa của chu kỳ bào tử được đưa ra dưới đây:
A. Bào tử:
Sự sinh bào tử bao gồm một chuỗi các sự kiện rất phức tạp trong sự biệt hóa tế bào (Hình 2.14). Nhiều thay đổi theo hướng di truyền trong tế bào tạo ra sự chuyển đổi từ tế bào ẩm, hoạt động trao đổi chất thành bào tử khô, không hoạt động về mặt chuyển hóa nhưng cực kỳ kháng thuốc.
Những thay đổi cấu trúc xảy ra trong các tế bào bào tử có thể được chia thành nhiều giai đoạn. Toàn bộ quá trình bào tử mất khoảng vài giờ. Các nghiên cứu di truyền đã chỉ ra rằng có tới 200 gen liên quan đến một quá trình sinh bào tử điển hình.
Trong quá trình sinh bào tử, quá trình tổng hợp một số protein liên quan đến hoạt động của tế bào sinh dưỡng chấm dứt và quá trình tổng hợp protein bào tử cụ thể bắt đầu. Những protein này được gọi là "protein bào tử hòa tan axit nhỏ" (SASPs).
Điều này được thực hiện bằng cách kích hoạt nhiều loại gen đặc hiệu bào tử bao gồm spo, ssp (mã hóa SASP) và nhiều gen khác để đáp ứng với điều kiện môi trường bất lợi. Các protein được mã hóa bởi các gen này xúc tác cho chuỗi các sự kiện thay đổi tế bào sinh dưỡng thành bào tử.
Các bào tử tăng khả năng chịu nhiệt của nó bằng một số cơ chế, chưa được giải thích rõ ràng. Tuy nhiên, rõ ràng là các lớp không thấm nước, chẳng hạn như vỏ bào tử và áo bào tử bảo vệ bào tử khỏi mọi thiệt hại vật lý.
Một yếu tố chính ảnh hưởng đến khả năng chịu nhiệt của bào tử là lượng và trạng thái của nước bên trong nó. Trong quá trình sinh bào tử, nguyên sinh chất bị giảm đến một thể tích tối thiểu, là kết quả của sự tích tụ Ca 2+, protein bào tử hòa tan axit nhỏ (SASPs) và tổng hợp axit dipicolinic, dẫn đến sự hình thành cấu trúc giống như gel.
Ở giai đoạn này, một vỏ não dày hình thành xung quanh lõi protoplast. Sự co thắt của vỏ não dẫn đến một nguyên sinh chất bị co rút, mất nước với hàm lượng nước chỉ bằng 10-30% so với tế bào sinh dưỡng. Hàm lượng nước của protoplast và nồng độ của SASP quyết định khả năng chịu nhiệt của bào tử.
Nếu bào tử có nồng độ thấp của SASP và hàm lượng nước cao, nó cho thấy khả năng chịu nhiệt thấp. Nếu nó có nồng độ cao của SASP và hàm lượng nước thấp, nó cho thấy khả năng chịu nhiệt cao. Nước di chuyển tự do trong và ngoài bào tử. Do đó, nó không phải là không thấm nước của áo bào tử, mà là vật liệu giống như gel trong nguyên sinh chất bào tử, không bao gồm nước.
B. Nảy mầm:
Một endospore có thể không hoạt động trong nhiều năm, nhưng có thể chuyển đổi trở lại một tế bào sinh dưỡng tương đối nhanh chóng. Quá trình này bao gồm ba bước; kích hoạt, nảy mầm và phát triển. "Kích hoạt" liên quan đến việc kích hoạt bào tử và làm cho nó sẵn sàng cho sự nảy mầm.
"Nảy mầm" liên quan đến việc mất độ khúc xạ siêu nhỏ của bào tử và mất khả năng chịu nhiệt và hóa chất. Các bào tử mất các thành phần canxi-dipicolinic và vỏ não. Hơn nữa, các SASP bị xuống cấp.
"Hết sức" liên quan đến sưng có thể nhìn thấy do sự hấp thụ nước cũng như tổng hợp DNA, RNA và protein mới. Tế bào nổi lên từ lớp bào tử bị hỏng và cuối cùng bắt đầu phân chia. Các tế bào sau đó vẫn còn trong tăng trưởng thực vật cho đến khi điều kiện môi trường thuận lợi.
Trong khi khử trùng một vật liệu để làm cho nó không có vi khuẩn, vi khuẩn có trong nó bị giết bằng phương pháp nào đó. Tuy nhiên, nếu vật liệu chứa các chất tạo bào tử, việc khử trùng sẽ trở nên khó khăn. Số lượng bào tử trong vật liệu càng nhiều thì ứng suất cần thiết để khử trùng nó càng cao.
Đối với các khuẩn lạc của vi khuẩn hình thành bào tử phát triển trên đĩa thạch hoặc xiên, khi tuổi vi khuẩn tăng lên, chúng hình thành bào tử do lão hóa, thiếu chất dinh dưỡng và tích lũy các chất chuyển hóa.