Nhiễm sắc thể Prokaryotic: Những lưu ý hữu ích về nhiễm sắc thể Prokaryotic
Dưới đây là những lưu ý của bạn về nhiễm sắc thể Prokaryotic!
Thông tin di truyền của một tế bào prokaryote được mang trong nhân tế bào trên cấu trúc gọi là nhiễm sắc thể của vi khuẩn. Nó bao gồm một phân tử DNA xoắn kép (deoxyribonucleic acid), không bao giờ liên kết với các protein cơ bản, và đã được chứng minh trong một số prokaryote là hình tròn.

Hình ảnh lịch sự: hangthebankers.com/wp-content/chromosome.jpg
Do đó, nhiễm sắc thể của vi khuẩn không tương đồng về cấu trúc với nhiễm sắc thể hạt nhân của tế bào nhân chuẩn, mà thay vào đó là DNA của tế bào cơ quan có trong ty thể và lục lạp. Có khả năng là một nhiễm sắc thể của vi khuẩn (viz., Một phân tử DNA rất dài) mang tất cả các thông tin di truyền cần thiết để xác định các tính chất cần thiết của tế bào prokaryote.
Plasmid: 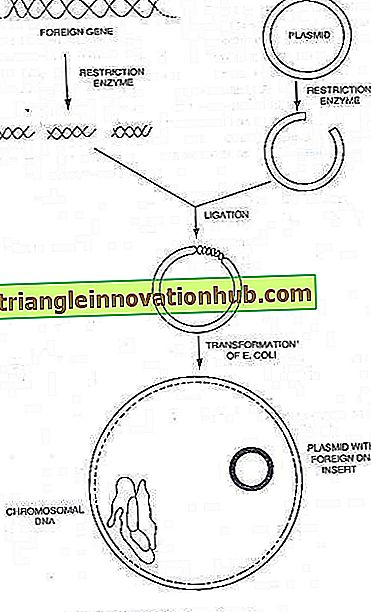

Nhiều vi khuẩn cũng có thể chứa các phân tử DNA hình tròn nhỏ, nhiễm sắc thể bổ sung có khả năng sao chép tự trị, được biết đến như là plasmid cho đến nay đã điều tra mang các yếu tố quyết định cho các đặc tính kiểu hình như kháng thuốc và các chất kháng khuẩn khác . Lượng DNA trong một plasmid là từ 0, 1 đến 5% trong số đó trong nhiễm sắc thể của vi khuẩn.
Điều này đã được phát hiện ở Escherichia coli rằng có hai loại giao phối và trong quá trình kết hợp, một đối tác chỉ đóng vai trò là người hiến tặng gen hoặc nam và người kia chỉ là người nhận gen hoặc nữ. Vì chức năng duy nhất của con đực là chuyển một số DNA của nó, nên nó không cần phải tồn tại, trong khi tế bào cái phải duy trì được khả năng, do đó, hợp tử có thể phát triển trong đó.
Điều này đã được phát hiện ra rằng sự ác tính ở vi khuẩn được xác định bởi một yếu tố di truyền khi vi khuẩn đực và cái kết hợp với nhau, mọi tế bào nữ được chuyển đổi thành con đực. Yếu tố di truyền chi phối tính chất di truyền của bệnh tật được gọi là yếu tố F (F = khả năng sinh sản), nó chỉ được truyền bởi tế bào trực tiếp tiếp xúc với tế bào. Năm 1952, Lederberg đã đặt ra thuật ngữ plasmid như một tên di truyền cho tất cả các yếu tố quyết định di truyền thêm nhiễm sắc thể, trong đó F là một ví dụ.
Hiện tại người ta đã biết rằng các plasmid của vi khuẩn là các phân tử DNA nhỏ, tròn mang gen để sao chép riêng. Trong nhiều trường hợp, chúng cũng mang các gen tạo ra các đặc tính mới trên tế bào chủ, chẳng hạn như kháng thuốc hoặc sản xuất độc tố. Nhiều plasmid mang gen chi phối quá trình liên hợp. Do đó, liên hợp là một cơ chế được áp đặt trên tế bào vi khuẩn bởi một plasmid, kết quả bình thường của nó là chuyển DNA plasmid.
Các loại plasmid:
Hầu hết các plasmid đã được phân loại trên cơ sở các thuộc tính của vật chủ. Do đó, có các yếu tố R (R = kháng) và các yếu tố COI (COI = colicinogeny) của vi khuẩn gram âm, các plasmid penicillinase của Staphylococcus aureus, các plasmid thoái hóa của Pseudomonas, plasmid phân hủy, v.v.
Tính chất của plasmid:
Cấu trúc của tất cả các plasmid đã biết bao gồm các phân tử DNA sợi tròn, kép. Một số trong số chúng có trọng lượng phân tử trong phạm vi 5 × I0 7 đến 7 × 10 7 . Một (một yếu tố R) có trọng lượng phân tử chỉ 1 × 10 7, và một số plasmid mật mã thậm chí còn nhỏ hơn. Vì số lượng DNA cần thiết để mã hóa cho một polypeptide trung bình có trọng lượng phân tử là 40.000 là khoảng 6 × 10 7, F 1 và các plasmid khác có kích thước tương tự có thể chứa tới 100 gen.
Biểu hiện gen ở Prokaryote:
Nhiễm sắc thể vi khuẩn:
Hiện tại người ta đã xác định rằng một nhiễm sắc thể của vi khuẩn luôn luôn tồn tại ở trung tâm của tế bào vi khuẩn không có khả năng phân chia phân bào và vi khuẩn và không được bao bọc trong màng nhân xác định. Một tế bào vi khuẩn trung bình chứa một phần nghìn hàm lượng DNA của một tế bào bạch đàn điển hình.
Nhiễm sắc thể của vi khuẩn là một sợi dài và mềm hình tròn của phân tử DNA liên kết với một số protein. Các tế bào vi khuẩn có thể phân chia rất nhanh. Sự phân chia tế bào được hoàn thành bằng cách nhân đôi tất cả các thành phần tế bào, sau đó phân vùng tế bào thành hai tế bào con. Bước đầu tiên trong phân chia tế bào là nhân đôi DNA.
Tái tổ hợp di truyền ở vi khuẩn:
Đây là một quá trình trong đó các vật liệu di truyền, chứa trong hai bộ gen riêng biệt, được kết hợp trong một đơn vị. Ở vi khuẩn, sự tái tổ hợp diễn ra bằng cách (1) biến đổi, (2) tải nạp và (3) liên hợp. Những hiện tượng này đã được thảo luận chi tiết trong các đoạn trước, dưới tiêu đề "sinh sản hữu tính ở vi khuẩn".

Cảm ứng và đàn áp:
Tiềm năng di truyền của virus bị hạn chế và về cơ bản nó sử dụng bộ máy chủ để nhân lên. Mặt khác, vi khuẩn có thể tổng hợp thành phần của chúng từ muối và đường đơn giản. Vi khuẩn, Escherichia coli có khoảng 2500 gen trên nhiễm sắc thể của nó. Vi khuẩn duy trì nền kinh tế nội bộ của nó chỉ bằng cách cho phép hoạt động của một số gen tại một thời điểm.
Điều này có nghĩa là các gen sẽ được bật hoặc tắt theo yêu cầu. Một bộ gen sẽ được bật khi cần thiết phải xử lý và chuyển hóa cơ chất mới. Khi bộ gen này được bật, các enzyme được tạo ra, giúp chuyển hóa cơ chất mới. Hiện tượng này được gọi là cảm ứng và các phân tử (chất chuyển hóa) gợi lên hiện tượng này là cảm ứng.
Mặt khác, khi một chất chuyển hóa cần thiết cho vi khuẩn được cung cấp vượt quá từ bên ngoài, vi khuẩn sẽ ức chế tạo ra nó và điều này ngăn chặn sự sản xuất thêm chất chuyển hóa của vi khuẩn. Do đó, các gen bất hoạt này được đặt tên là có thể ức chế và hiện tượng này được gọi là đàn áp hoặc ức chế phản hồi.
Tuy nhiên, một số gen liên tục được thể hiện để chăm sóc hoạt động bình thường của tế bào như glycolysis. Những gen này được gọi là cấu thành và các enzyme do chúng tạo ra được gọi là enzyme cấu thành, chẳng hạn như dehydrogenase.
Khái niệm toán tử:
Francois Jacob và Jacques Monod (1961), khi làm việc tại Viện Pasteur, Paris, đã đề xuất rằng các con đường trao đổi chất được quy định như một đơn vị. Họ đã nghiên cứu sự thay đổi bản chất của các enzyme của E. coli khi được cung cấp các loại đường khác nhau. Họ đã tìm thấy, khi đường được thêm vào các nền văn hóa của E. coli, nó tạo ra ba loại enzyme cần thiết để phá vỡ đường sữa thành glucose và galactose.
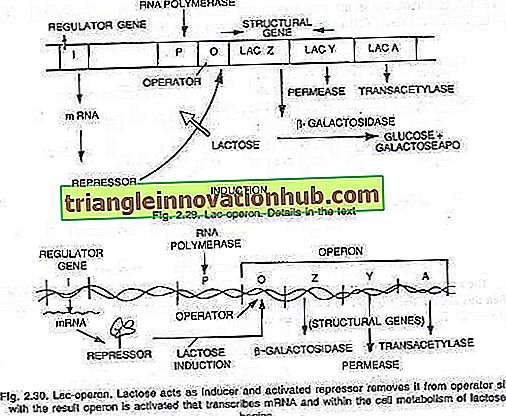
Các enzyme mới được tổng hợp này là- (i) p-galactosidase, (ii) permease và (iii) transacetylase. Theo họ, sự tổng hợp của ba enzyme này được kiểm soát bởi một đoạn DNA dài gọi là operon phân chia thành một vùng vận hành O và ba gen cấu trúc Z, Y và A. Hoạt động của các gen cấu trúc được điều khiển bởi vị trí của người vận hành với sự giúp đỡ của một protein ức chế được tạo ra bởi tác động của gen 'i' được gọi là gen điều hòa.
Các gen được biểu thị hay không biểu hiện phụ thuộc vào việc bật hay tắt công cụ vận hành. Khi bật công tắc, ba gen được RNA polymerase phiên mã thành một đoạn RNA thông tin duy nhất bao trùm cả ba gen. Mỗi gen cấu trúc thường được gọi là cistron và mRNA dài được phiên mã bao gồm tất cả các cistron được gọi là polycistronic.
Việc bật hoặc tắt công tắc vận hành được thực hiện bởi một protein được gọi là repressor. Khi protein này liên kết với người vận hành (O) và chặn nó, công tắc sẽ bị tắt và ba gen (Z, Y, A) không được biểu thị, (xem hình 2.28).
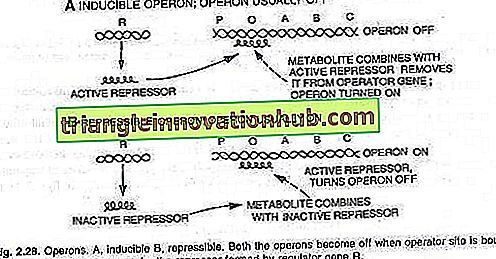
Lạc-operon:
Đây là một ví dụ về operon cảm ứng. Một vài phân tử đường sữa được thêm vào tế bào nhờ hoạt động của enzyme permease, một lượng nhỏ được tìm thấy ngay cả trong điều kiện bị ức chế. Một vài phân tử này sau đó được chuyển đổi thành một dạng hoạt động của đường sữa, liên kết với chất ức chế (sản phẩm gen i) và do đó, chất ức chế không thể liên kết với người vận hành nữa.
Khi người vận hành không có chất ức chế RNA polymerase, anh ta bắt đầu phiên mã operon bằng cách liên kết vị trí quảng cáo P. Messenger RNA tương ứng với cả ba enzyme được dịch mã để tạo ra ba enzyme - P-galactosidase, permease và transacetylase.
Với việc sản xuất ba enzyme chuyển hóa đường sữa này bắt đầu. Quá trình tổng hợp enzyme được tiếp tục trừ khi và cho đến khi tất cả các phân tử lactose được tiêu thụ bởi tế bào. Khi các phân tử cuối cùng của đường sữa, liên kết với chất ức chế cũng được tiêu thụ, chất ức chế không hoạt động sẽ hoạt động và liên kết với vị trí vận hành (O) để tắt operon như bình thường, (xem hình 2.29 và 2.30).
Tryptophan-operon:
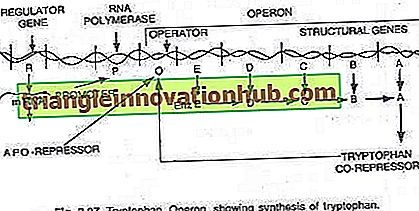
Trong trường hợp này, operon thường bật để sao chép và dịch mã để tổng hợp các enzyme cần thiết cho quá trình tổng hợp một chất chuyển hóa nhất định của tế bào. Tuy nhiên, operon có thể được tắt khi tế bào không yêu cầu chất chuyển hóa hoặc chất chuyển hóa đã được sản xuất dư thừa. Toán hạng tryptophan bao gồm năm gen (trp E, D, C, B và A) mã hóa cho năm enzyme (En z E, D, C, B và A) xúc tác cho quá trình tổng hợp tryptophan (một axit amin) và do đó tạo thành một con đường đồng hóa. Sự hiện diện của tryptophan phục vụ cho việc kìm hãm sự tổng hợp của các enzyme chịu trách nhiệm sản xuất nó, (xem hình 2.27 và 2.31).
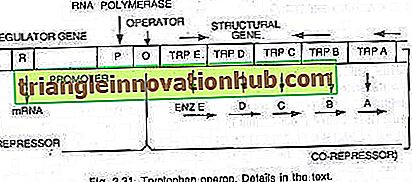
Năm gen cấu trúc mã hóa cho năm enzyme này được điều chỉnh bởi một công tắc vận hành duy nhất. Ở đây, protein repressor được tạo ra bởi các gen điều hòa không tự liên kết với người vận hành. Tuy nhiên, khi tryptophan được thêm vào, nó liên kết với chất ức chế và được gọi là chất ức chế. Phức hợp repressor-corepressor liên kết với toán tử và chặn biểu thức của operon này.
Cấu trúc của operon tryptophan ít nhiều giống với cấu trúc của operon lac, nhưng có một biến thể chức năng. Trong trường hợp này, sản phẩm gen R tương đương với gen i của operon lac. Sản phẩm gen R tạo ra protein mà bản thân nó không thể liên kết với người vận hành. Điều này được gọi là apo-repressor.
Với sự có mặt của tryptophan, một đồng ức chế, bộ ức chế chức năng được hình thành, hiện đang liên kết với toán tử và ngăn chặn sự phiên mã của operon và sản xuất tryptophan. Ở đây mô hình operon giải thích cả hiện tượng cảm ứng và đàn áp trong Procaryote.